Neuron mechanisms for defining receptive fields and behavioural recommendation weights
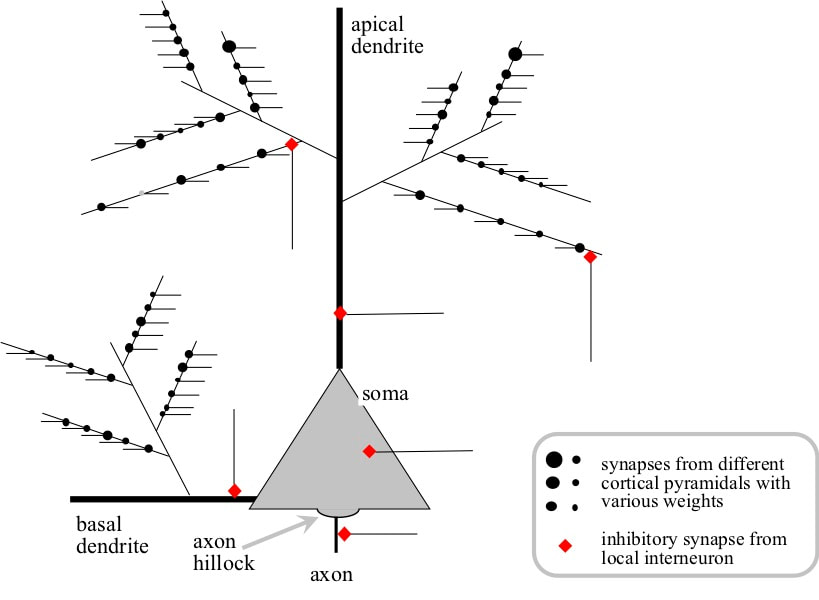
Neurons are cells, and like all other human cells they have a compact body surrounded by a cell wall or membrane. Inside the membrane are a number of structures including a nucleus with the DNA genetic information. In neurons this compact body is called the soma, but unlike most cells outside the brain neurons have a number of bulges in the cell membrane that extend long distances away from the soma. One such extension is the axon. This axon branches extensively to carry any outputs from the neuron to many target neurons. Each output is a 100 millivolt electrical spike called an action potential. The other type of extension is the dendrite. A neuron can have multiple dendrites, and most inputs from other neurons arrive on the dendrites. The place where an incoming axon contacts a dendrite is called a synapse. Synapses define the type and strength of the effect of an input on the target neuron.
PYRAMIDAL NEURONS IN THE CORTEX DEFINE AND DETECT RECEPTIVE FIELDS Detection of a receptive field at some point in time is indicated by an action potential spike launched along the axon. Receptive fields of neurons in layer V of the cortex have direct behavioural recommendation strengths as a result of connectivity on to neurons in the striatum of the basal ganglia. Other receptive fields have implicit recommendation strengths because they target layer V neurons, directly or via other neurons.
The dendrites of a pyramidal neuron branch extensively to form a tree like structure. A pyramidal neuron has one apical dendrite that emerges on the opposite side of the cell body from the axon, and multiple basal dendrites that emerge from the cell body in various directions perpendicular to the line defined by the axon and apical dendrite.
The final “twigs” on the dendritic tree are called terminal branches. There are about 140 terminal branches on an average pyramidal, and most of the inputs from other pyramidal neurons arrive on these terminal branches.
When an incoming action potential arrives at a synapse, it injects an electrical charge called a postsynaptic potential into the terminal branch. The magnitude of this postsynaptic potential is called the synaptic weight, and is different for different synapses.
Cortical Pyramidal Neuron
Each terminal branch detects a group of very similar conditions. Each of these conditions is defined by a combination of synapses that has sufficient total synaptic weight that if action potentials arrive at all the synapses in the group within a few milliseconds of each other the terminal branch injects potential deeper into the dendrite. A dendrite detects a group of more complex conditions. Each dendritic condition is defined by a combination of terminal branches that, if all the branches inject potential into the dendrite within a short period of time, will result in the dendrite injecting potential into the body of the neuron. The receptive field of the neuron is defined by different groups of dendritic conditions that result in the neuron generating an output action potential.
The pyramidal neuron receptive field is thus defined by a large group of complex combinations of inputs from other pyramidals. Each combination has sufficient total synaptic weight to drive an output. A receptive field can therefore be changed by addition of synapses and/or by changes to synaptic weights. However, because receptive fields are associated with behavioural recommendation strengths, such changes are limited and carefully managed to minimize side effects on the validity of those recommendation strengths.
SYNAPTIC MECHANISMS FOR RECEPTIVE FIELD CHANGES Pyramidal neurons use a neurotransmitter called glutamate to communicate across synapses. The synaptic weights of glutamatergic synapses from one pyramidal on to another define receptive fields.
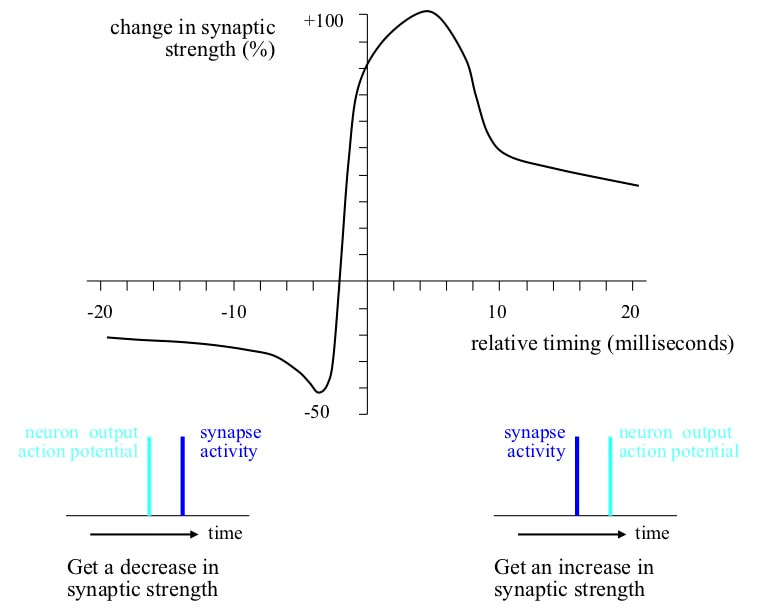
Use of temporal correlation information The key mechanism for synaptic weight changes uses the relative timing between arrival of a spike at a synapse and firing of the target neuron. If a spike arrives at a synapse and shortly afterwards the neuron fires, the weight of the synapse increases. However, if a neuron fires and shortly afterwards a spike arrives at the synapse the synaptic weight decreases, although the decreases are smaller than the increases.
This effect of relative timing is the primary mechanism for receptive field changes. If a spike from some source neuron arrives just before the target neuron fires, the relative timing indicates that the receptive field of the source neuron is probably relevant to future detections of the target neuron receptive field. If the source neuron spike arrives after the receptive field detection by the target, the source neuron receptive field is probably irrelevant. In other words, the mechanism supports definition of receptive fields on the basis of temporal correlations.
Use of reward feedback Reward feedback cannot be used to define receptive fields, but can be used to confirm changes made strictly on the basis of temporal correlations. Synaptic weight changes made on the basis of relative spike timing decay back to their original values over a period of about an hour. If there is reward feedback indicating that recently detected receptive fields were important for determining significant behaviours, the decay is blocked and the changes are retained long term.
Threshold changes If a pyramidal neuron fires very frequently, detections of its receptive field are not discriminating between situations in which its recommendation strengths are very relevant and those in which they are less relevant. In this situation, the effective threshold for firing is increased, without changing the type of circumstances in which it detects its receptive field.
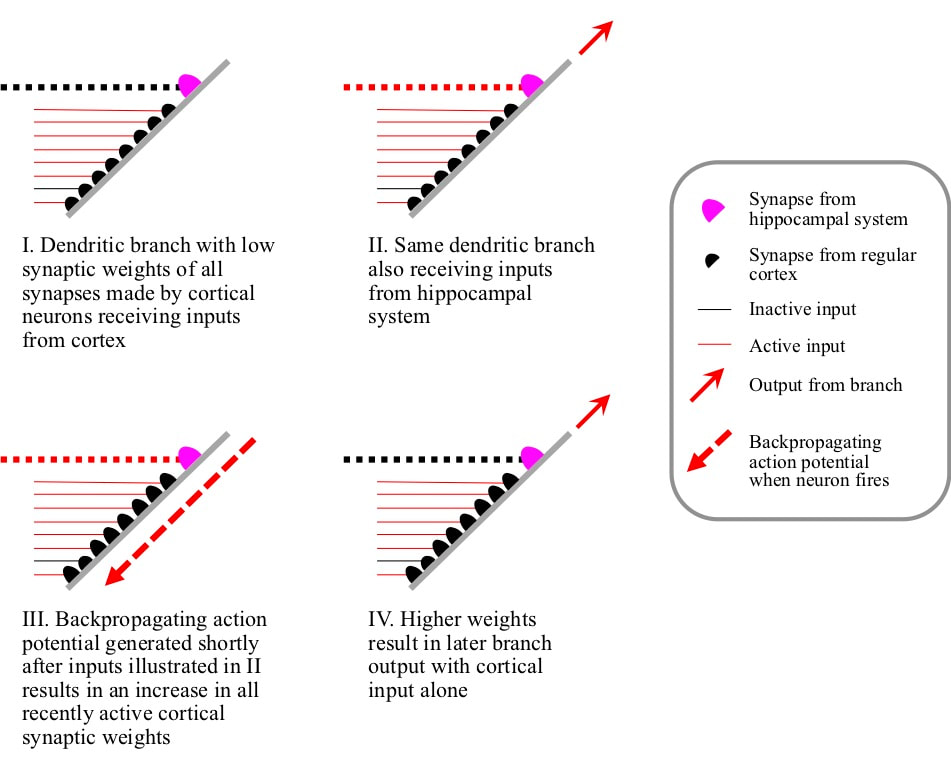
Role of the hippocampal system When the total number of receptive field expansions is too small, the hippocampal system amplifies the regular temporal correlation mechanism to drive additional receptive field expansions. Outputs from the hippocampal system establish synapses on the branches of cortical pyramidal neurons. If a branch is not detecting any condition on the basis of the weights of its synapses that are receiving inputs from cortical pyramidal neurons, an input from the hippocampal system may be sufficient to make the total postsynaptic potential large enough for the branch to inject potential into its dendrite. If the neuron fires shortly afterwards, the synaptic weights of all the synapses on the branch that had active cortical inputs will increase. These increases mean that if exactly the same combination of inputs occurred in the future, there would be enough postsynaptic potential for the branch to inject potential into its dendrite without a hippocampal input. In other words, a new condition has been defined on the branch.
The hippocampal mechanism can add rather more different conditions. For example, some synapses made by cortical pyramidals on to other cortical pyramidals have zero weight. A branch could have a number of zero weight or silent synapses, and might never detect any conditions in the absence of hippocampal input. However, if with the aid of hippocampal input the branch injects potential deeper into the dendrite and a little later the neuron fires, silent synapses that recently received an input can gain weight.
MEDIUM SPINY NEURONS IN THE STRIATUM OF THE BASAL GANGLIA DEFINE AND INTEGRATE RECOMMENDATION STRENGTHS Outputs from layer V pyramidal neurons target medium spiny neurons in the striatum of the basal ganglia. Each medium spiny neuron corresponds with one behaviour, and the strength of a pyramidal neuron synapse on to the medium spiny neuron is the recommendation weight of the source cortical receptive field in favour of that behaviour.
The dendrites of a medium spiny neuron emerge in many directions from the soma to form a rough star shape. Synapses from many different cortical pyramidal neurons target these dendrites. When an incoming action potential arrives at a synapse, it injects an electrical charge called a postsynaptic potential into the terminal branch. The magnitude of this postsynaptic potential is called the synaptic weight, and is different for different synapses.
One pyramidal neuron makes a very small number of synapses on to one medium spiny neuron, probably just one synapse. Inputs at many synapses are required to fire the neuron. If a lot of inputs arrive close together at the same synapse, the total effect is much less that the synaptic weight multiplied by the number of inputs. However, if inputs arrive at the same time at multiple synapses, the total effect is much larger than the total of the individual synaptic weights.
Each synaptic weight is the recommendation weight of the pyramidal neuron receptive field in favour of the behaviour corresponding with the medium spiny neuron. A behaviour can only be selected if it is recommended by many different cortical receptive fields.
SYNAPTIC MECHANISMS FOR RECOMMENDATION WEIGHT CHANGES The synaptic weights of glutamatergic synapses from a pyramidal neuron on to a medium spiny neuron in the striatum of the basal ganglia define recommendation weights. Just as for connections between pyramidal neurons, synapses are isolated on spines so that synaptic weights can be changed independently.
When a behaviour is selected, the medium spiny neurons in the striatum firing most strongly are those corresponding with that behaviour. Again as for connections between pyramidal neurons, the weights of any synapses that received inputs just before the neuron fired are increased.
If a positive reward is indicated after the behaviour, the weight increases are made permanent. However, in the case of the medium spiny neurons, if a negative reward is indicated, the recent weight increases are reversed and the weight set at a lower level than initially. Hence rewards act to make long terms changes to recommendation weights.
BRAIN INFORMATION MECHANISMS AT THE NEURON LEVEL The key information mechanisms for the brain are receptive field (or condition) definition/detection and behavioural recommendation definition/integration. At the neuron level, receptive field detection is indicated by the firing of cortical pyramidal neurons, and receptive fields are defined by glutamatergic synapses on to those pyramidal neurons. Behavioural recommendation strengths are defined by the strengths of glutamatergic synapses made by pyramidal neurons on to medium spiny neurons in the striatum of the basal ganglia. The integration of the recommendation strengths in performed by the medium spiny neurons.