The role of the cerebellum is to speed up the execution of often used sequences of behaviours
The ability to carry out behaviours rapidly has considerable natural selection advantages. In the brain, the implementation of some behaviours can be considerably speeded up by the cerebellum. The cerebellum is a large structure with two hemispheres located behind the brainstem at the back of the brain below the cortical hemispheres. The cerebellar hemispheres have an outer sheet of neurons surrounding a body of white matter (i.e. axons). Buried inside the cerebellar hemispheres is a set of nuclei called the cerebellar nuclei. The importance of the cerebellar role is indicated by the fact that it has about 60% of the neurons in the brain.
REGULAR MECHANISMS FOR CONTROL OF BEHAVIOUR In order to determine behaviour, the brain defines large numbers of conditions that are combinations, or combinations of combinations etc., of the available information. This available information includes raw sensory inputs from the environment, but also includes large amounts of information about the state of the body and of the brain itself. The conditions are recorded on pyramidal neurons in the cortex, and are called the receptive fields of those neurons. At each point in time, any conditions that are currently present are detected, and detections are indicated by the firing of the corresponding pyramidal neurons.
The basal ganglia gets inputs from many of the cortical pyramidal neurons, and interprets the firing of any such neuron as a recommendation in favour of a range of different behaviours, each recommendation having an individual weight. Processing of this information by the basal ganglia identifies and implements the behaviour with the largest total recommendation weight across all currently detected receptive fields. Other receptive fields detected by the cortex a little while after the behaviour has been carried out can have recommendation strengths in favour of changing the weights that recommended any recent behaviours. This allows reward type information to adjust recommendation weights over time.
SEQUENCES OF BEHAVIOURS There are many situations in which a sequence of behaviours often needs to be carried out in the same order. When climbing stairs, a sequence of muscle movements is required to move one foot up to the next stair, another sequence to move the other foot, then the same sequences repeat. Those sequences are used whenever there is a need to climb stairs. Speech is generated by the coordinated movement of around 100 different muscles in the face, tongue and voice box. If a word or phrase is often spoken, the sequence of muscle movements that generate the word or phrase is often carried out in the same order. A cognitive process like recalling the memory of some event is implemented by a sequence of behaviours that release information between specific cortical areas. This type of sequence may also be performed many times to recall memories of different events.
ADVANTAGES OF SHIFTING CONTROL OF BEHAVIOUR SEQUENCES TO THE CEREBELLUM When the basal ganglia controls the performance of a sequence of behaviours, receptive fields detected when one behaviour is completed contribute much of the recommendation strength in favour of initiating the next behaviour. Hence under basal ganglia control, selection of the next behaviour generally occurs only after the preceding behaviour has been completed. Once selected, time is required for the communication of the selection out to the muscles or other structures that carry out the behaviour. All this severely limits the speed with which sequences of behaviours can be carried out.
However, such sequences of behaviours can be recorded in the cerebellum. Once recorded, a sequence is selected once as a whole by the basal ganglia, then executed by the cerebellum with no further reference to the basal ganglia.
Neurons in the cerebellum develop receptive fields that correspond with all the different circumstances in which one behaviour in a sequence should be initiated. These circumstances can include information about the completion of behaviours much earlier in the sequence, and the execution of the sequence can therefore proceed much faster. Detection of such a cerebellar receptive field can be viewed as a command to carry out the corresponding behaviour, in contrast with detection of a cortical receptive field, which is only a recommendation in favour of a range of behaviours.
The use of behaviourally unambiguous information (i.e. commands) in the cerebellum means that the ability to learn is severely restricted. The timing of the behaviours in the sequence can be speeded up, but no changes can be made to the identity of any of the behaviours or the order in which they are carried out. To make changes of these types, behavioural control must revert to the basal ganglia.
If someone decides to pick up some object, the basal ganglia selects the behaviour of reaching for the object. The cerebellum then performs the sequence of arm, hand and finger muscle movements to carry out the action. A patient with damage to the cerebellum affecting just movements on the right side has some difficulties with this process. Reaching with the left arm proceeds smoothly and feels unconscious. Reaching with the right arm has to be thought out step by step.
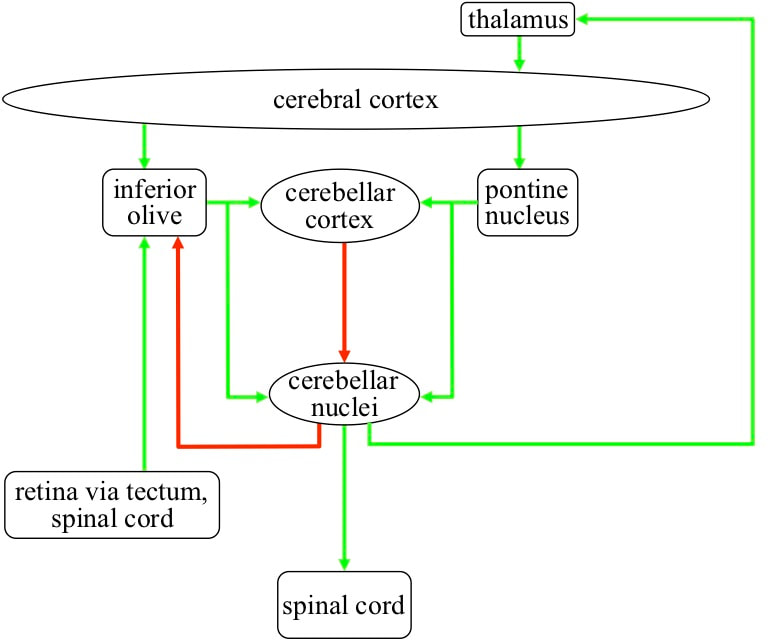
STRUCTURE AND CONNECTIVITY OF THE CEREBELLAR SYSTEM The cerebellar system has a number of components that together carry out its role. These components include the cerebellar cortex and the cerebellar nuclei, plus two nuclei located in the brainstem, the pontine nucleus and the inferior olive.
The cerebellar cortex has two layers of neurons. The inner layer contains huge numbers of very small neurons called granule cells. The layer above it contains the central bodies of very large neurons called Purkinje cells. Above the neuron layers is a layer that contains the dendrites of the Punkinje cells. Axons from the granule cells go up to this outer layer, and each Purkinje gets inputs from of the order of 100 thousand different granule cells. In the cerebellar nuclei there are two types of neuron, one excitatory, the other inhibitory. The Purkinje cells target both these types of cerebellar nuclei neurons.
Thick bundles of axons, called the cerebral peduncles, carry outputs from pyramidal neurons in the cerebral cortex to the cerebellar system. These axon bundles have about 20 million axons and mainly target the pontine nucleus. The pontine nucleus has perhaps 10 million neurons, each of which gets thousands of inputs from the cerebral cortex. Outputs from the pontine nucleus target the granule cells, and each granule cell gets multiple inputs from just three or four pontine neurons. Then as mentioned earlier, each Purkinje gets inputs from about 100 thousand granule cells. A significant number of granule cell inputs are needed to cause a Purkinje to fire, but much less than the total number of granule cell inputs. So a Purkinje neuron fires in response to any one of a very large number of different combinations of granule cell inputs, and each granule cell fires in response to a combination of three or four extremely complex combinations of cortical receptive fields. In addition, the pontine nucleus outputs also target the neurons in the cerebellar nuclei.
The inferior olive gets inputs from the cortex, and also from the spinal cord, and from the retina of the eye by a fast route that bypasses the regular route into the brain via the thalamus. In addition the inferior olive gets inputs from the inhibitive neurons in the cerebellar nuclei. Outputs from inferior olive neurons target Purkinje cells. A Purkinje gets inputs from just one inferior olive neuron. The axon of that neuron wraps itself around the dendrite of the Purkinje, making hundreds of synapses.
The only outputs from the cerebellar system come from the excitatory neurons in the cerebellar nuclei. These neurons target the spinal cord to drive motor behaviours, and also target the thalamus to drive cognitive behaviours.
ZONES WITHIN THE CEREBELLAR SYSTEM The connectivity between neurons in the cerebellar system is organized in zones. Each zone contains about a hundred Purkinje neurons, and about the same numbers of inferior olive neurons, excitatory neurons in the cerebellar nuclei, and inhibitory neurons in the cerebellar nuclei. Connectivity between these neurons is just within the zone to which they belong.
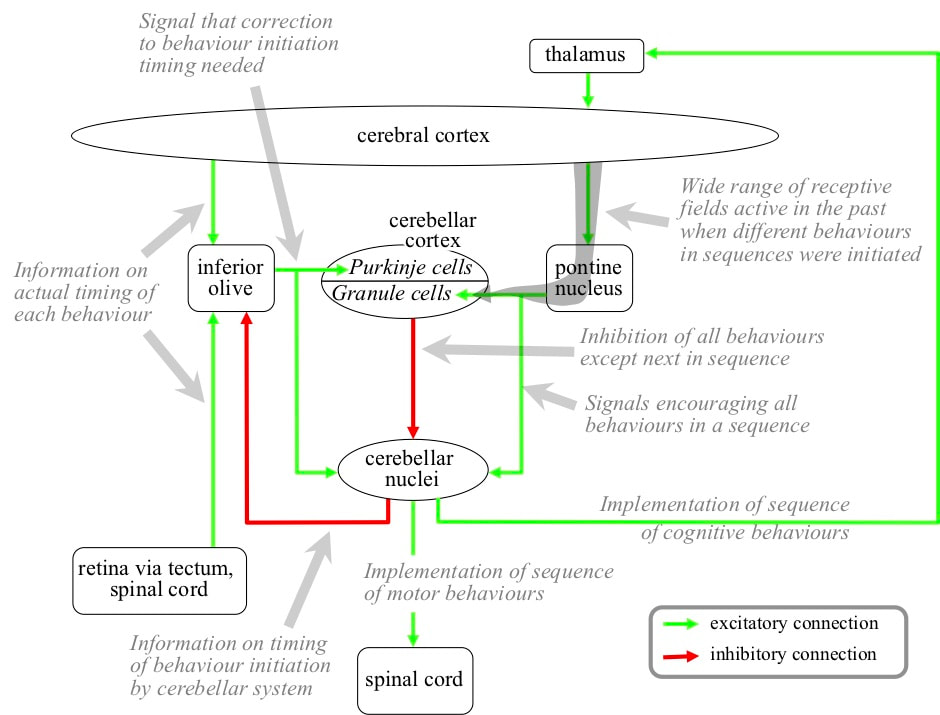
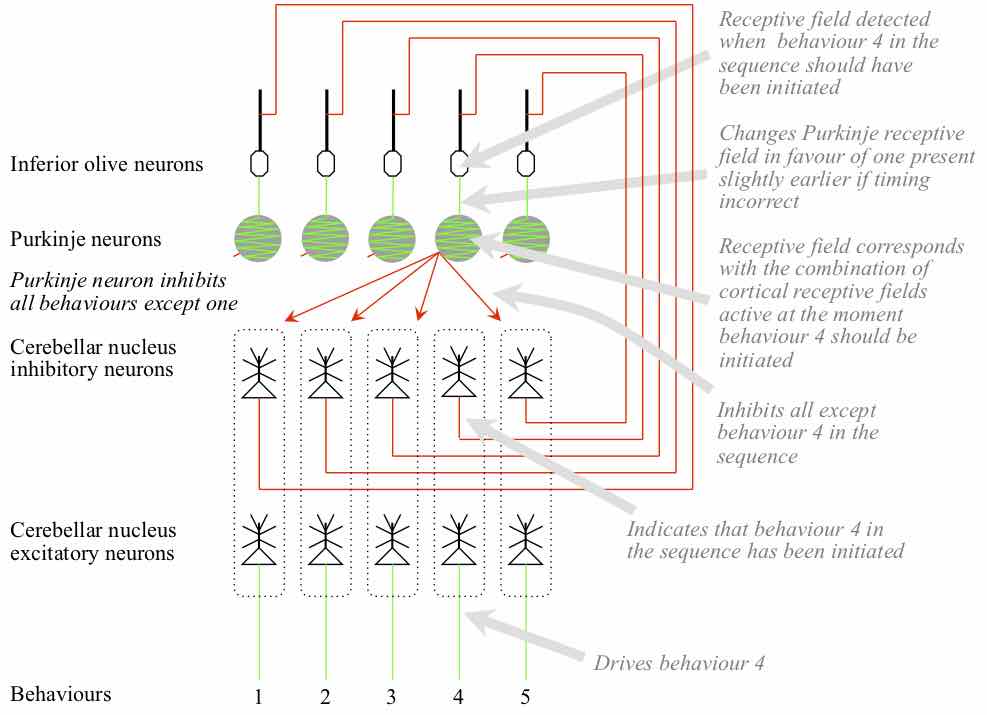
INFORMATION PROCESSES IN THE CEREBELLAR SYSTEM A cerebellar zone corresponds with one behaviour sequence. Each Purkinje in a zone corresponds with a different behaviour in the sequence. The receptive field initially developed by a Purkinje corresponds with whatever combinations of cortical pyramidal neurons happen to be active at the moment the behaviour was initiated when the basal ganglia was controlling behaviour selection.
The cerebellar neurons in a zone also correspond with a different behaviour in the sequence. The excitatory neuron targets the neurons in the spinal cord and/or thalamus that were active when the corresponding behaviour was initiated under the control of the basal ganglia. Hence the excitatory neurons in the cerebellar nuclei drive their corresponding behaviour. The inhibitory neuron targets the inferior olive neuron that in turn targets the Purkinje corresponding with the same behaviour. When the inferior olive neuron fires soon after its target Purkinje, it increases the synaptic weights of some of the granule cell inputs to its target Purkinje. The weights that are increased are those of granule cells that fired ~100 milliseconds before the Purkinje cell itself fired. Hence the effect of the inferior olive firing is to shift the Purkinje receptive field towards a field that would have been detected a little earlier.
A Purkinje cell targets all the cerebellar nuclei neurons in its zone, except those corresponding with its own behaviour. Purkinje outputs are inhibitory, hence the firing of a Purkinje inhibits all the behaviours in a sequence except its corresponding behaviour.
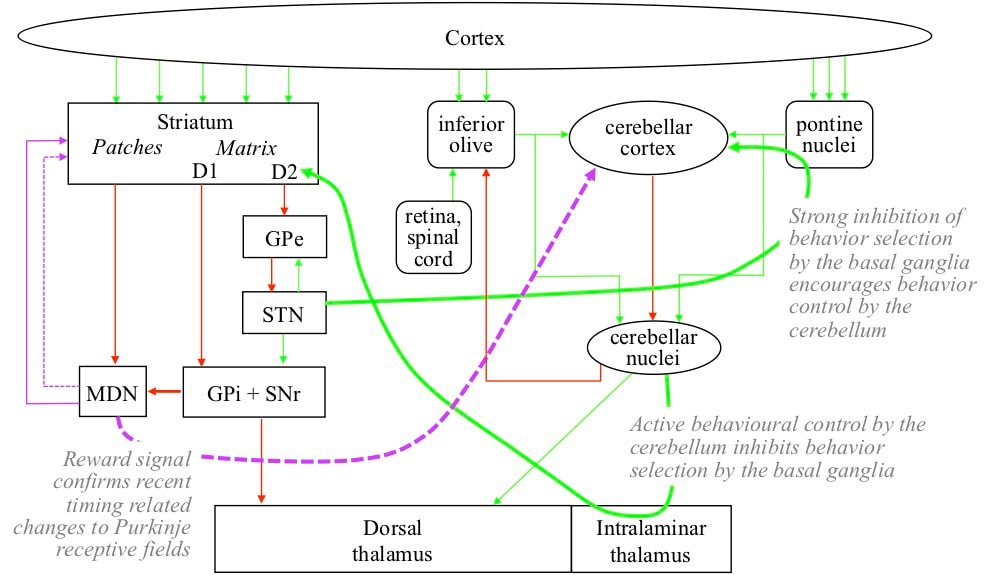
EXECUTION OF A BEHAVIOUR SEQUENCE In circumstances in which the behaviour sequence is appropriate, a range of cortical receptive field detections detected within those circumstances recommend its performance. If there is sufficient recommendation strength in favour of the sequence, the basal ganglia release a wide range of cortical receptive field detections to the cerebellar system. Connectivity from the cerebellum to the basal ganglia then discourages behaviour control by the basal ganglia by increasing activity of D2 striatal neurons at the start of the indirect path, via intralaminar nuclei in the thalamus. Connectivity from the basal ganglia to the cerebellum encourages behaviour control by the cerebellum because the subthalamic nucleus (STN) in the indirect path increases activity in the pontine nucleus.
The increased level of input from the pontine nucleus encourages all the excitatory cerebellar nuclei neurons in the zone to fire. The receptive fields of the Purkinje cells are detected in the sequence in which their behaviours are performed. The firing of a Purkinje inhibits all the excitatory cerebellar nuclei neurons except the one corresponding with its behaviour. Hence the behaviours are executed in the appropriate sequence.
The inhibitory cerebellar nuclei neurons in the zone also fire in the sequence controlled by the Purkinje cells. These cerebellar nuclei neurons target inferior olive neurons that also correspond with the different behaviours in the sequence. The inferior olive neurons get input from the cortex, spinal cord, and eyes derived from the actual implementation of the behaviour sequence. Their receptive fields are detected at the moment their corresponding behaviour should be implemented. If the cerebellar nuclei initiate the behaviour at that moment, firing of the inferior olive is inhibited. If the behaviour is initiated later, the inferior olive neuron fires, targeting the Purkinje that drives initiation of the behaviour. The effect of the inferior olive firing is to shift the receptive field of the Purkinje so that it will be detected slightly earlier in the future. Hence the effect is that the corresponding behaviour will be initiated slightly earlier. Over time, the Purkinje field evolves towards whatever combinations of cortical receptive fields were active in the past when the behaviour needed to be initiated.
SIGNIFICANCE OF THE CEREBELLUM A behaviour is learned by cortical receptive fields acquiring recommendation strengths in favour of the behaviour in the striatum of the basal ganglia. Once learned, behaviour selection by the basal ganglia takes a significant amount of time. For sequences of behaviours that have been fully learned, this time can be avoided and the performance of the sequence speeded up by transfer of control to the cerebellum. A zone in the cerebellum learns a sequence and can drive execution of the sequence in quick succession.
Because the basal ganglia does not control the individual behaviours, no changes can be made to the identity of the behaviours or their order. To make such changes, control must revert to the basal ganglia. The only learning possible in the cerebellum is reduction of the time between behaviours in an established sequence.
A very wide range of behaviour sequences are controlled by the cerebellum in this fashion. Examples include the sequences of muscle movements that constantly maintain our balance, and the sequences of muscle movements that keep our limbs in position when we are sitting still to prevent our bodies collapsing like wet rags. The sequences of eye movements when we scan different types of objects can also be controlled by the cerebellum.
Movement sequences such as those required for walking, running, climbing stairs or riding a bicycle are also under cerebellar control. One interesting aspect of sequences like those needed to ride a bicycle is that often the sequences learned initially in the basal ganglia do not work properly because they cannot be carried out fast enough to maintain, for example, balance. The difficulty in learning such skills results from the need for control to shift back and forth between the cerebellum and basal ganglia.
Sequences often used in cognition are also coded in the cerebellum. One simple example is the sequences of muscle movements used to speak often used words and phrases. A more complex example is the sequences of releases of information between different cortical areas needed to carry out often used cognitive processes like recalling the memory of events.