Sleep is defined as a period each day when an animal goes into a physically passive state. During sleep, the brain is much less sensitive to sensory inputs. Activities like seeking food or caring for offspring are suspended. Because sensory processing is partially suspended, the animal is very vulnerable to environmental threats such as from predators. The need for sleep increases steadily while awake, and only goes back to zero after an adequate period of sleep, which is longer after a period of deprivation.
Such sleep appears to be ubiquitous in any living thing with a brain, including humans, other mammals, birds, reptiles, fish and insects. Sleep deprivation results in cognitive problems – from memory, mood and attention problems in humans to reduced precision in the dances that signal good pollen locations in honeybees.
Although body movements are limited, the brain is not passive. On the contrary it is highly active, although the pattern of activity is qualitatively different from when awake. In humans there are two phases of sleep, labelled rapid eye movement (REM) sleep after the phenomenon by which it was first identified, and non-REM (NREM) sleep. If someone is wakened during REM sleep they report vivid visual dreams, while any reported dreaming when wakened from NREM sleep is more like thinking. There is a tendency for dreaming in the early part of the night to have some resemblances to recent experiences, while dreaming later in the night resembles more distant experience. Experiences in which there was novelty or in which rewards or strong emotions occurred tend to feature more often. However, if there are no external interruptions, generally very little is recalled about dreams after waking up.
THE PRIMARY PROBLEM ADDRESSED BY SLEEP Why is sleep ubiquitous in all animals with brains, given the high vulnerability of the animal during sleep? Why is there such vigorous brain activity during sleep? To understand what is going on, recall that there are two general types of information processes in the brain, condition (or receptive field) definition/detection and behavioural recommendation definition/integration.
The practical problem is that any changes must occur in time periods of milliseconds to seconds, and this does not provide enough time to grow additional synapses if required. Some provisional synapses must therefore be created in advance to allow later learning. Again, the brain does not “know” in advance what provisional synapses will be needed. Such synapses could be created randomly, but this would mean either excessive numbers of useless synapses or the risk that some necessary learning could not occur. The brain addresses this problem by placing a bias on the random creation of synapses.
The only available guide to future experience is past experience, with recent experience somewhat more likely to be relevant. To identify synapses likely to be useful in the future, the brain performs a fast approximate rerun of fragments of past experience, the most recent experiences occurring earliest in the sleep period. This fast partial rerun is labelled vivid dreaming. Synapses are created between neurons that are often active at the same time during this rerun. If two neurons have often been active at the same time in the past, there is a chance that a connection between them could be useful. A connection between neurons that were never active at similar times is unlikely to be relevant.
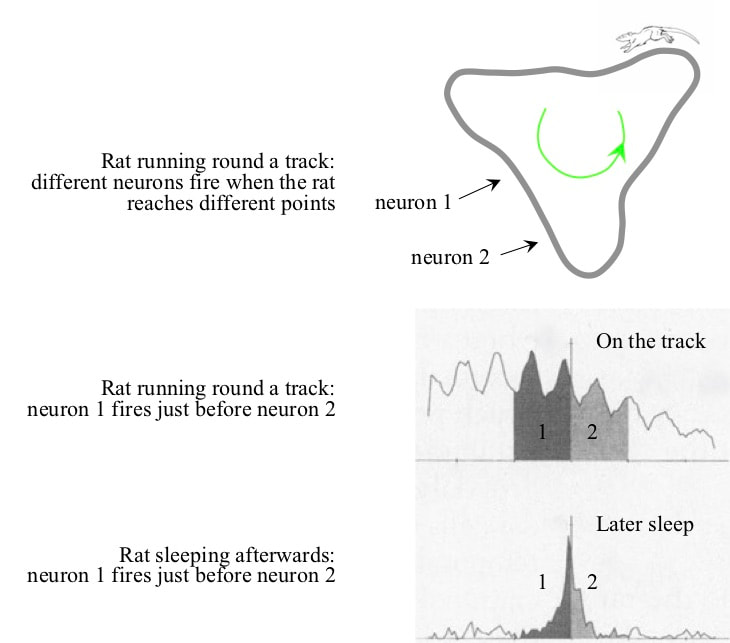
In sleep, hippocampal neurons fire in the same order as during waking experience. Some hippocampal neurons fire when the brain is in one specific location. Hence when a rat runs around a track, neurons fire in a particular order. During sleep, the neurons fire in the same order, but about seven times faster
Such a fast rerun would interfere with the processing of current experience, and the brain is therefore taken off-line. The rerun has some general similarities with actual waking experience but is often likely to result in cross connections between unrelated experiences. If the brain recalls some fragment of dreaming, the memory can therefore often be bizarre.
A key role of sleep is therefore to create additional synapses that are likely to be useful for recording relevant information during experiences in the next waking period. The synapses most likely to be relevant are identified by fast reruns of fragments of the past which are experienced as sometimes bizarre dreams. Axon, dendrite and synapse growth occurs mainly during NREM sleep.
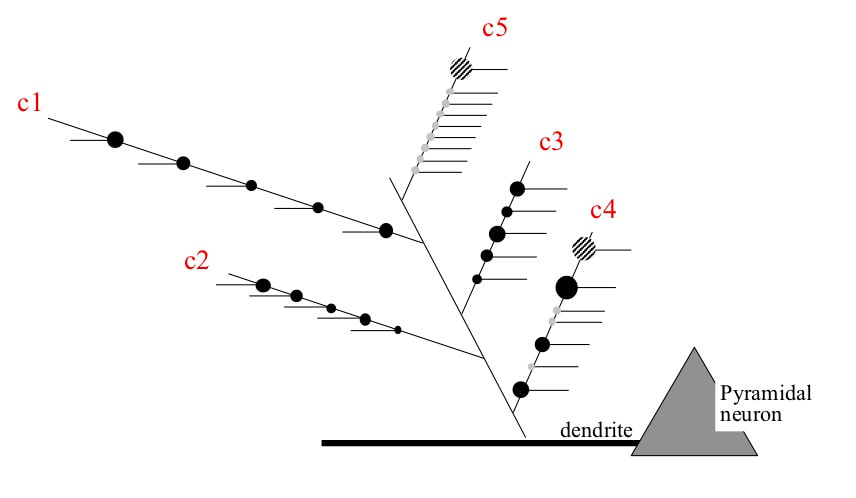
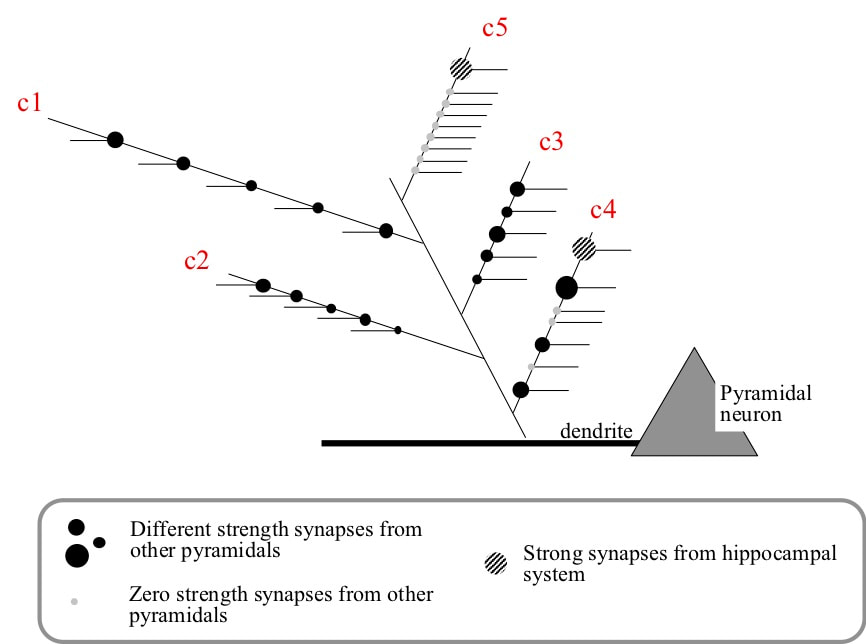
Framework for changing the receptive field of a cortical pyramidal neuron. The receptive field is made up of conditions instantiated on terminal branches of dendrites. Each condition is defined by a combination of synapses from other pyramidal neurons. Provisional synapses with zero weights are created during sleep. Small changes to the receptive field can occur by increasing the synaptic weights of existing inputs to a branch (e.g. c1, c2, c3). Large changes could occur by the hippocampal system driving addition of weight to a branch made up of zero weight provisional synapses (e.g. c5). Intermediate changes can occur by the hippocampal system driving increases to synaptic weights on branches made up of a mix of regular and zero weight synapses (e.g. c4)
If for some period of waking experience, very little active processing by the brain is needed, recall of recent experience could be used to guide growth of new connectivity without interfering with current processing. Hence when learning a new skill, taking passive breaks between learning episodes can enhance learning. During such breaks, reruns of neuron activity during skill learning can be observed, and learning is more rapid when such breaks are taken.
ALTERNATIVE THEORIES FOR THE ROLE OF SLEEP Sleep could have a number of non-cognitive roles, from immobilization to reduce exposure to predators to various chemical maintenance tasks. Such roles could well be significant, taking advantage of the sleep period, but the vigorous and atypical activity of the brain during sleep suggests that the primary reason is support for some brain function. One popular alternative theory to the one outlined above is that during waking, the information that is the basis for future memory recalls is recorded in some temporary store such as the hippocampus, and during sleep the information is transferred to some long term storage such as the cortex. Such transfer is sometimes called consolidation.
The origin of this curious idea was the observation of catastrophic interference in artificial neural networks, in which new learning completely erased earlier learning. It was suggested that to avoid this problem, when new information was recorded, it was put in a temporary store. In subsequent sleep, large parts of both recent and more remote experiences are reactivated in such a way that they can be combined in a way that minimizes interference.
In detail, the mechanism to run recent and past experiences with high precision so that they could be reliably moved to a different storage location would be very complex and therefore biologically implausible. In the brain the interference problem is addressed by very careful management of cortical changes by the hippocampal system. The reruns to identify appropriate provisional connectivity have no need for high precision reproduction of past experiences.
SIGNIFICANCE OF SLEEP TO COGNITION To learn appropriate behaviours, the brain needs to define receptive fields in the cortex and recommendation strengths of receptive fields in the striatum of the basal ganglia. The receptive fields of cortical pyramidal neurons are defined by combinations of the receptive fields of other pyramidals. There are immense numbers of receptive fields that could be defined on this basis, and with limited resources the brain must identify those more likely to be useful. Many different receptive fields could potentially contribute recommendation strength to the MSNs associated with any one behaviour, and again the brain must favour those likely to be useful. The use of information about correlations in neuron activity during past experiences is the best available information about relevant future connections that is available to the brain.
During sleep, neuron resources are configured in such a way that they are more effective for the type of learning that will be needed in the immediate future. This configuration process uses past experience as the best available guide to the form of future experiences, with the recent past a bit more likely to be relevant than the more distant past. To identify the neuron connectivity most likely to be useful in future experiences, the brain performs a rapid rerun of fragments of past experiences and establishes connectivity between neurons active at the same time during this rerun. Because learning is especially important for novel experiences and experiences in which there were strong rewards (positive or negative) the content of the rerun is biased in favour of fragments of such experiences.
If a brain needs to learn new material or a new skill, on first exposure learning is somewhat less effective. After a period of sleep, new connectivity has been created, and learning will generally be more effective. Sleep thus configures neuron resources likely to be useful for learning in the next wake period, increasing the efficiency with which the neuron resources are used.