How the cortex and basal ganglia together select brain behaviour
A pyramidal neuron in the cortex detects the presence of one or more of a group of similar conditions. This group is called the receptive field of the neuron. Unlike in a computer system, where detection of a condition is interpreted as a command to perform a behaviour, in the brain a condition detection is just a recommendation in favour of many different behaviours, each recommendation having its own weight.
Pyramidal neurons in layer V of the cortex target medium spiny neurons in a part of the basal ganglia called the striatum. Each medium spiny neuron corresponds with one behaviour or type of behaviour. The synaptic weight of the connection from the pyramidal neuron on to the medium spiny neuron is the recommendation weight of the receptive field in favour of the behaviour corresponding with the medium spiny neuron.
What are the behaviours that can be selected?. There are two general types managed by the basal ganglia. One type targets the cortex, releasing selected subsets of currently detected conditions into and/or out of specific cortical areas. Release of a selected subset of current sensory inputs into a primary sensory area is called an attention behaviour. Release of selected receptive field detections between two cortical areas is a cognitive behaviour. Release of selected receptive field detections from the motor cortex to drive muscles is a motor behaviour. The other general type of behaviour is a reward behaviour. Reward behaviours increase or decrease the recommendation weights that selected recent behaviours. These reward behaviors target the striatum.
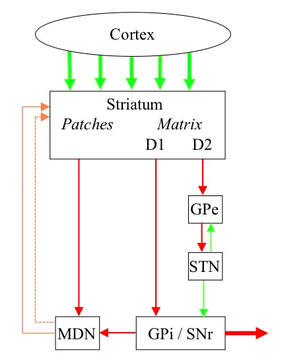
The striatum of the basal ganglia is made up of two intermingled structures called the matrix and the patches. Medium spiny neurons in the matrix correspond with different release behaviours. Medium spiny neurons in the patches correspond with different reward behaviours.
STRATEGIC, TACTICAL, SPECIFIC AND DETAILED BEHAVIOURS One way to classify behaviours is along a spectrum from strategic through tactical and specific to very detailed. For example, think about a musician playing a concert. The selection of the time and place of the concert is at the strategic end of the spectrum. The selection of the pieces of music to be played is more tactical. The finger movements to play the instrument are very detailed.
Different cortical areas define receptive fields that are combinations of inputs from three or four other areas. Areas therefore detect receptive fields in different ranges of complexity. Some ranges of complexity are more effective for discriminating between circumstances in which different strategic behaviours are appropriate. Receptive fields in areas with this complexity therefore tend to recommend different strategic behaviours. Other areas are most effective for recommending more tactical behaviours and so on.
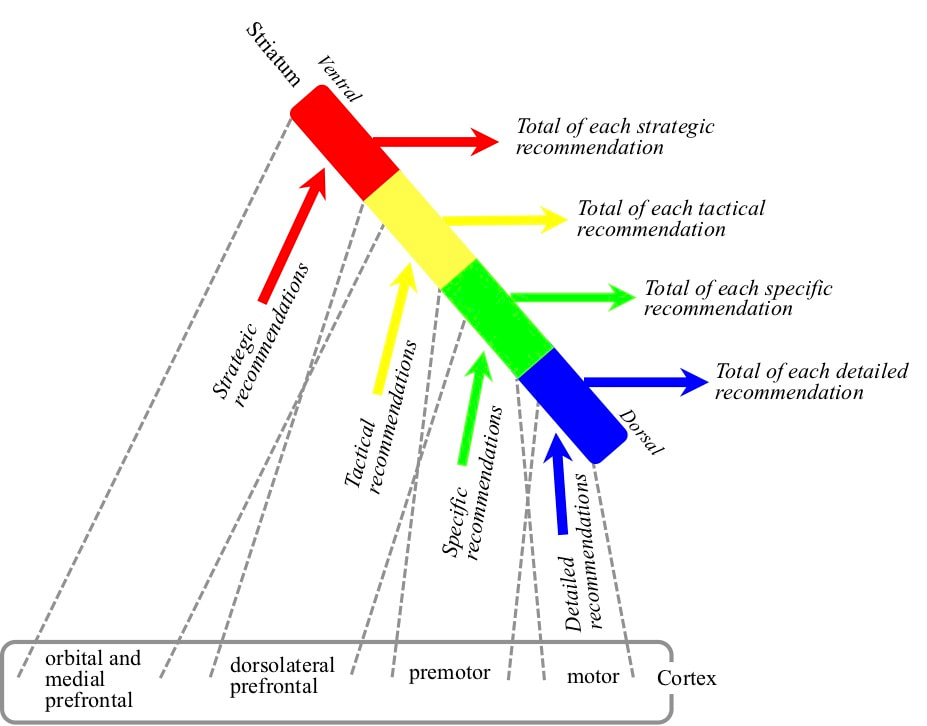
Different cortical areas are more effective for recommending different general types of behaviour from strategic to very detailed muscle movements. Each receptive field detection recommends a wide range of different behaviours of the general type into the striatum in the basal ganglia. The striatum determines the total weight in favour of each specific behaviour of each general type. Different parts of the striatum from ventral to basal determine the total weights for different general types of behaviour from strategic to detailed.
The striatum is a long structure extending from a ventral end to a dorsal end. Strategic behaviours are selected at the ventral end and detailed behaviours at the dorsal end. Areas in the orbital and medial prefrontal cortex are most effective for recommending strategic behaviours but also to a lesser degree tactical behaviours. Areas in the dorsolateral prefrontal cortex are most effective for recommending more tactical behaviours but also to a certain degree specific behaviours. Areas in the premotor area are most effective for recommending specific behaviours but also to a certain degree detailed behaviours. Areas in the motor cortex are effective for recommending detailed behaviours. The targetting of the striatum by the cortex reflects the higher recommendation effectiveness of different areas.
I mentioned earlier that these behaviours are implemented by release of receptive field detections. How do such releases implement more strategic behaviours? The answer is that the receptive fields most strongly recommending the selected strategic behaviour are released to areas effective for recommending more tactical behaviours. This tends to result in the receptive fields detected in the target areas having recommendation strengths in favour of different tactical behaviours of the selected strategic type. Once a tactical behaviour is selected, it is implemented by release of the receptive field detections that most strongly recommended it to areas effective for recommending more specific behaviours and so on.
SELECTING ONE BEHAVIOUR BUT NOT MORE THAN ONE The behaviour selection processes in the basal ganglia have to achieve a couple of results. Firstly, it is important to select a behaviour most of the time, but to avoid selecting two incompatible behaviours at the same time, such as trying to move an arm in two directions at once. Secondly, whatever behaviour is selected must be implemented.
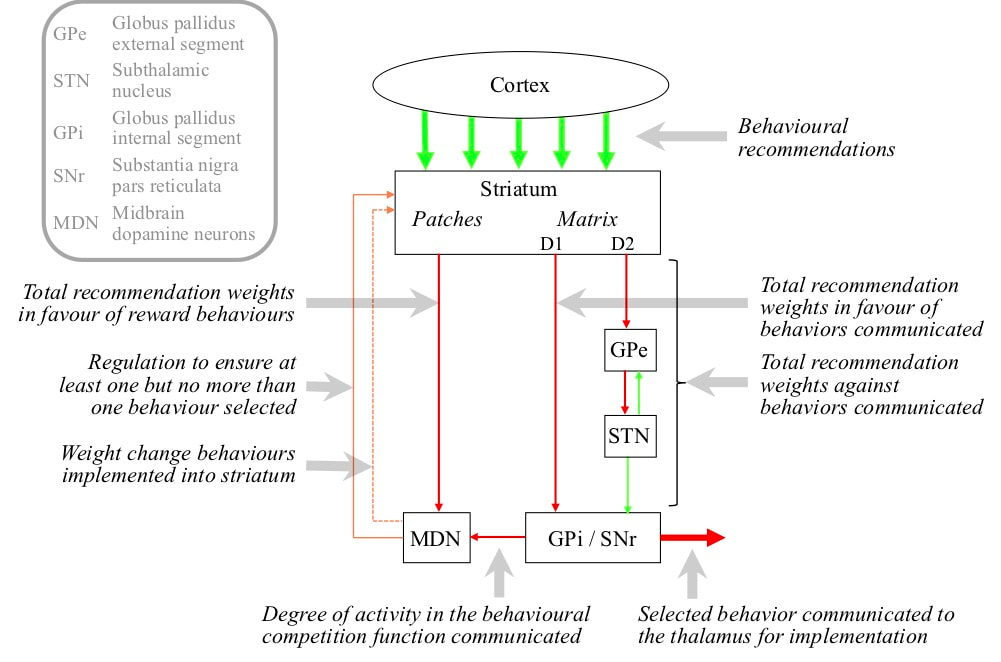
The connectivity of the basal ganglia delivers these results. A neuron in a structure called the GPi/SNr corresponds with one behaviour, and constantly fires in a way that inhibits that behaviour. The matrix of the striatum contains two kinds of medium spiny neurons labelled D1 and D2. The firing of D1 neurons corresponds with the total recommendation strength of their current cortical inputs in favour of their corresponding behaviour, and they directly target GPi/SNr neurons corresponding with the same behaviour. D1 neuron firing inhibits that GPi/SNr neiron and therefore encourage the behaviour. The firing of D2 neurons corresponds with the total recommendation strength against any behaviour except their corresponding behaviour. They target GPi/SNr neurons corresponding with any behaviour except their own via an indirect path through two structures called the GPe and STN. Via this path, D2 neuron firing excites GPi/SNr, and therefore discourage any other behaviours. There is therefore a competition within the striatum and GPi/SNr that selects the most strongly recommended behaviour.
If the total activity in the GPi/SNr is high, the implication is that all behaviours are blocked and none is being selected. If the total activity in the GPi/SNr is low, the implication is that more than one behaviour is being selected. This total activity is communicated to another structure called the midbrain dopamine neurons or MDN. If GPi/SNr activity is too high, indicating that no behaviour being selected, MDN activity increases in such a way that the background dopamine in the striatum increases. This increases D1 firing relative to D2 and increases the chance of a behaviour being selected. On the other hand, If GPi/SNr activity is too low, indicating that multiple behaviours being selected, MDN activity decreases and the background dopamine in the striatum increases. This decreases D1 firing relative to D2 and reduces the chance of a behaviour being selected.
At a more detailed level, the dopaminergic neurons that regulate the striatum fire steadily at about four spikes per second. The GPi/SNr acts by changing the number of dopaminergic firing in this mode.
STRATEGIC, TACTICAL, SPECIFIC AND DETAILED REWARD BEHAVIOURS Recommendation weights must be adjusted depending on whether the consequences of the selected behaviour were positive or negative. Some cortical areas define receptive fields effective for discriminating between circumstances in which positive or negative rewards are appropriate. These receptive fields have recommendation weights in favour of rewards. Pyramidal neurons in these areas target medium spiny neurons in the patches of the striatum. These medium spiny neurons correspond with reward behaviours.
Receptive fields effective for rewarding strategic behaviours are somewhat different from those effective for rewarding tactical behaviours and so on. In the musician example used earlier, the presence of a large crowd at the concert could reward the strategic behaviour of selecting the time and place of the concert. Applause after each piece of music and at the end of the concert could reward the tactical behaviour of selecting the pieces. Receptive fields detected within hearing the music as it was generated or seeing the reactions of the audience could reward finger movements.
If there is sufficient recommendation strength in favour of a reward, the corresponding medium spiny neuron fires. If the reward is positive, this firing triggers the firing of dopaminergic neurons in the MDN that target back to the striatum. This dopaminergic neuron firing is a burst of spikes, about ten in half a second, that is quite different from the steady firing at four spikes per second regulating current behaviour selection. The matrix neurons corresponding with the recently selected behaviour will have fired strongly. Such firing temporarily increases the weights of any synapses that recently received an input from the cortex. If a burst dopamine input is received soon afterwards, the temporary increase is made permanent. In other words, the recommendation weights of the receptive fields that recommended the behaviour are increased. If the reward is negative, the effect is to temporarily reduce the number of dopaminergic neurons firing steadily. This reduction reduces the weights of any synapses that recently received an input from the cortex, reducing the recommendation weights of the receptive fields that recommended the recent behaviour.
REWARDING REWARD BEHAVIOURS Cortical inputs to medium spiny neurons in the striatal patches have recommendation weights in favour of the behaviours of rewarding recent behaviours. But how are those recommendation weights in favour of rewards themselves determined? In other words, how are reward behaviours rewarded? Once again the musician example can provide some clues. Think about how individual finger movements to generate notes are rewarded. When the musician was learning to play, a teacher’s words or expression indicated approval, and receptive fields detected within those teacher responses had recommendation strengths in favour of rewarding recent behaviours, including finger movements. Receptive fields defined within hearing the quality of the notes were detected at the same time as the teacher response fields, and these auditory fields therefore acquired recommendation strengths in favour of rewards. The question is, how could the recommendation weights of these rewarding rewards fields be further improved?
After playing a piece of music, there might be strong applause. This is a tactical reward signal, but would not be appropriate for rewarding every individual finger movement made while playing, some may have been better than others. However, the applause does indicate that on the whole that notes were good. The implication is that the way in which the notes are being rewarded is working well. So such applause could be appropriate for rewarding the way individual finger movements are being rewarded. In other words, the behaviour of rewarding a tactical behaviour could also reward the behaviour of rewarding more detailed behaviours.
In general, the behaviour of rewarding a more strategic behaviour could also reward the behaviour of rewarding tactical behaviours. The behaviour of rewarding a more tactical behaviour could also reward the behaviour of rewarding more detailed behaviours, and so on.
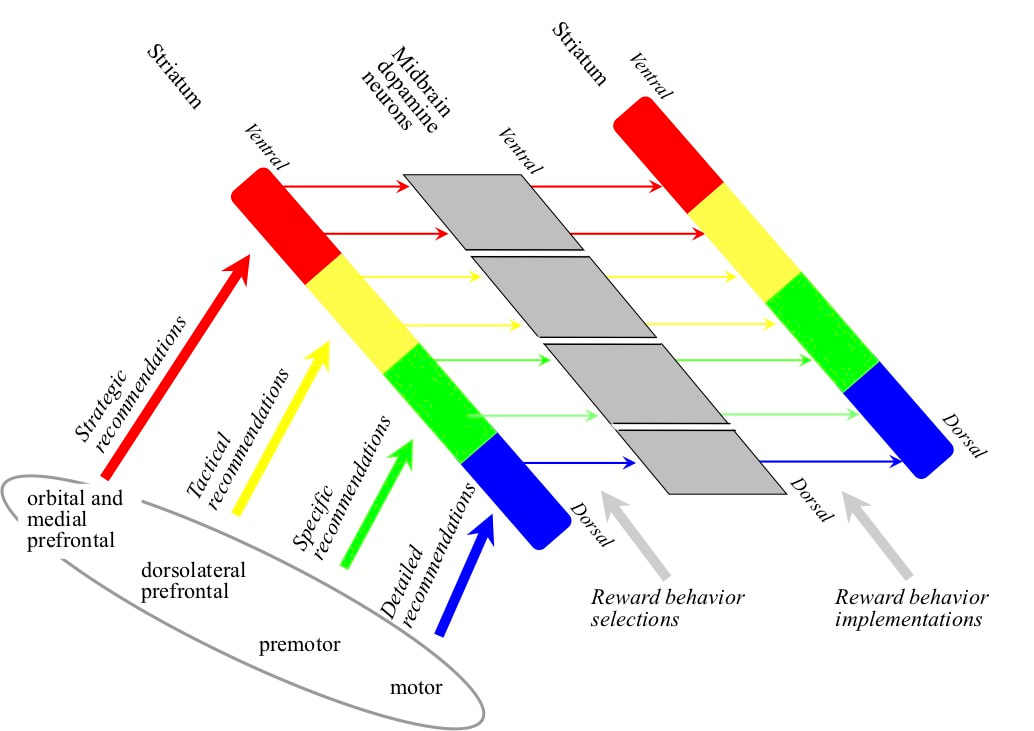
Both the striatum and the midbrain dopamine neurons are arranged from a more ventral region to a more dorsal region. The total recommendation strengths in favour of rewarding behaviours are determined in the striatum, the more strategic at the ventral end, the more detailed at the dorsal end. Outputs from the striatum communicating selected reward behaviours target the midbrain dopamine neurons. The midbrain dopamine neurons implement reward behaviours by outputs back to the striatum that change recommendation weights. However, there is a connectivity offset in the pattern of midbrain dopamine neuron outputs back to the striatum. The most ventral part of the midbrain dopamine neurons that is targetted by the most ventral part of the striatum actually targets the most ventral part of the striatum plus a portion of the less ventral striatum. This pattern carries on all across the two structures with each region of the midbrain dopamine neurons targetting the region of the striatum from which it received inputs plus a somewhat more dorsal striatal region. This offset pattern implements the rewarding of reward behaviours. The behaviour of rewarding a more strategic behaviour also rewards the behaviour of rewarding more tactical behaviours and so on.
INFORMATION PROCESSING IN THE BRAIN There are two general types of processes in the brain. Conditions are defined and detected in the cortex. Behavioural recommendation strengths are defined and used in the basal ganglia, using current cortical condition detections. The most complex information processing is performed by the cortex, but the basal ganglia are essential to interpret the results of that information