The roles of the hippocampus and cortical columns in managing changes to cortical receptive fields
Pyramidal neurons in the cortex define and detect receptive fields. A receptive field is defined by a group of conditions, and is detected if one of the group is present in its current inputs. Detection of the receptive field is associated with a range of recommendations in favour of different behaviours, each recommendation having its own individual weight. Receptive fields are defined heuristically from experience, and will therefore change over time. The problem this presents is that if a receptive field is changed, the change will generally make all the associated recommendation strengths less valid. If a receptive field is detected in somewhat different circumstances, some of the behaviours it recommends may be less appropriate in those circumstances. Alternatively, the field may no longer be detected in circumstances in which its recommendations are appropriate. The implication is that changes to cortical receptive fields must be very carefully managed to minimize undesirable side effects on behaviours.
WHAT KIND OF CHANGES CAN BE MADE TO CORTICAL RECEPTIVE FIELDS? Given that receptive field changes risk jeopardizing the integrity of all the recommendation strengths associated with the field, what kinds of changes are the least risky? If the changed receptive field is still detected in all the circumstances in which it was detected in the past, the risk is minimized. So the best strategy is a slight increase in the range of circumstances in which the receptive field is detected. Such increases are called receptive field expansions. Decreases have much higher risk, and tend to be more rare.
WHEN SHOULD CORTICAL RECEPTIVE FIELDS BE CHANGED? Given the risks involved, what would be a good reason to make changes to receptive fields? One receptive field recommends many behaviours, and the selected behaviour at each point in time is the one with the largest recommendation strength across all currently detected receptive fields. To ensure selection of an appropriate behaviour, a reasonably wide range of recommendations needs to be available. If only a few receptive fields are being detected, the range is limited. Hence if too few receptive fields are being detected, some receptive fields must be expanded so that they are also detected and add their recommendation strengths. In general, relatively few receptive fields will be detected if the circumstances are novel, so more expansions must occur in novel situations.
WHICH CORTICAL RECEPTIVE FIELDS SHOULD BE CHANGED? For two reasons, changes should be as small as possible. One reason is that there will be less disruption to the integrity of existing recommendation strengths. The second reason is that if a receptive field is already close to being detected there must be some degree of resemblance to the current situation, and relevant recommendation strengths are more likely. Hence in novel situations, relatively few receptive fields will be detected, and undetected receptive fields that are the closest to being detected must be slightly expanded until they are detected.
IDENTIFYING THE RECEPTIVE FIELDS TO BE CHANGED A big problem is, how can the brain identify the receptive fields that are the closest to being detected? Such identification requires comparisons between the internal states of receptive fields that are not currently being detected. Directly comparing the internal states of all the pyramidal neurons in the cortex and identifying which were closest to detection would be extremely difficult.
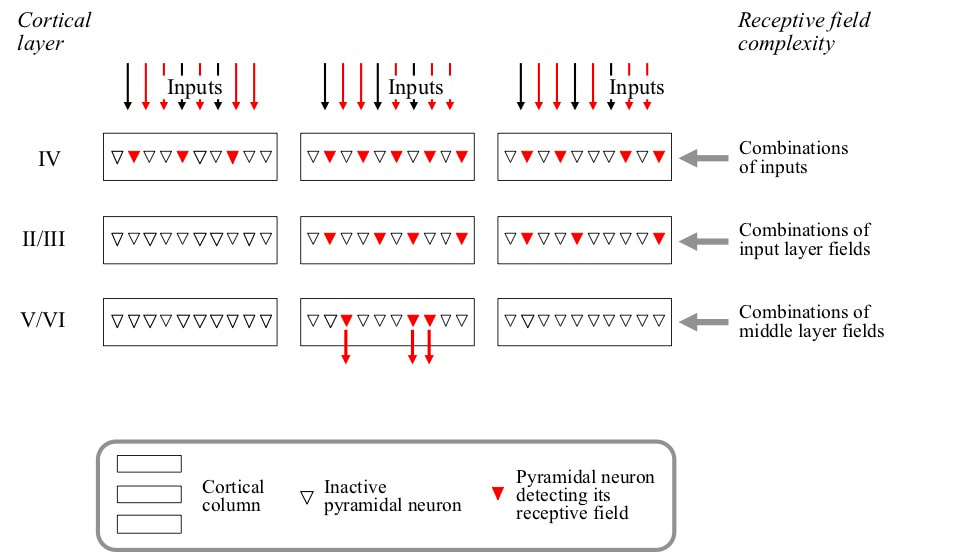
The columns observed in the cortex are a key part of the way the brain solves this problem. Cortical columns are groups of about 10 thousand pyramidal neurons arranged into a number of layers. Typically there are five layers labelled II, III, IV, V and VI. There is a pronounced pattern of connectivity across the layers of a column. External inputs arrive in layer IV. Outputs from layer IV go to layers II and III, and outputs from layers II/III go to layers V and VI. Layers V and VI generate outputs from the column.
All the neurons in a column have generally similar receptive fields. However, the receptive fields in layers II/III are defined by combinations of receptive fields in layer IV, and receptive fields in layers V/VI are defined by combinations of layers II/III receptive fields. Hence layers II/III are in connectivity terms the middle layers, and there is a gradual increase in receptive field complexity going IV –› II/III –› V/VI. The implication is that although the receptive fields in all layers are generally similar, they become more more specific going IV –› II/III –› V/VI, in the sense that they will be detected in fewer circumstances.
If receptive fields are detected in layers V/VI, the receptive field of the column is detected. However, if there is no activity in layers V/VI but strong internal activity in layers II/III, the receptive field is much closer to being detected than if there is weak or no internal activity. The cortical columns that are closest to being detected can therefore be identified on this basis, and receptive fields of pyramidal neurons of those columns targetted for expansion if expansions are needed.
There is another criterion that is useful for identifying the most appropriate columns for expansion. Some columns will be already detecting their receptive field when expansions are needed. There may be a number of the columns not detecting their receptive fields but having strong internal activity. If some of those columns expanded their receptive fields in the past at the same time as some of the columns already detecting their fields, that temporally correlated past expansions would make current expansion somewhat more likely to be appropriate.
Hence in novel situations, the best candidates for receptive field expansion are columns with strong layer II/III activity and no layer V/VI activity that expanded their receptive fields in the past at the same time as some columns already detecting their fields.
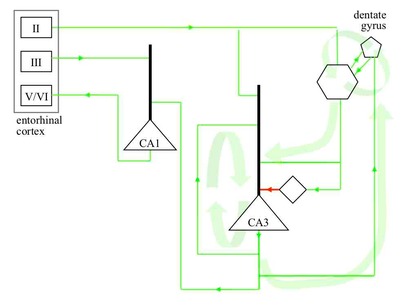
CHANGE MANAGEMENT BY THE HIPPOCAMPAL SYSTEM At each point in time, the hippocampal system identifies the appropriate cortical receptive fields for expansion, and sends signals to drive those expansions. Within the hippocampal system there is the hippocampus proper and three adjacent cortical areas, the perirhinal, parahippocampal and entorhinal cortices. The system also includes nuclei belonging to a number of subcortical structures, connected with the hippocampus proper by a thick axon bundle called the fornix.
The associated cortical areas get inputs from layers II/III of columns in all cortical areas except the primary sensory areas, and define receptive fields corresponding with groups of columns that expanded their receptive fields at similar times in the past. Layer II/III outputs from monomodal cortical areas go to perirhinal and parahippocampal cortices. Layer II/III outputs from polymodal cortices and the perirhinal and parahippocampal cortices go to the entorhinal cortex. Layer II/III outputs from the entorhinal cortex go to the hippocampus proper. Within the hippocampus proper there is a competition process to determine the most appropriate cortical columns for receptive field expansion, and outputs go back through the associated cortical areas to drive the expansions. In the course of this process, the receptive fields in those cortices are updated to reflect the current expansions. The primary sensory areas are excluded because the high degree to which receptive fields in these areas are defined under genetic control makes external change management unnecessary.
The hippocampus proper is made up of a number of structures, including two regions called CA1 and CA3 with pyramidal neurons like those in the cortex, and the dentate gyrus with two types of neurons, granule cells and mossy cells, both types excitatory like the pyramidal neurons.
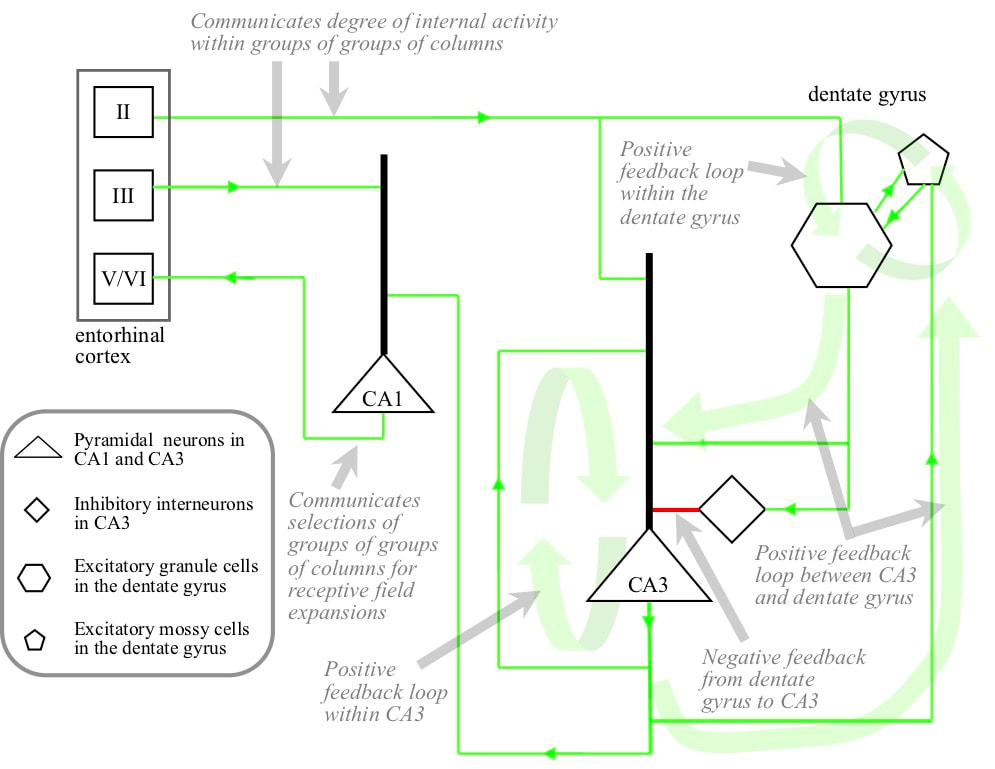
Layer II/III outputs from the entorhinal cortex go to CA1, CA3 and to granule cells in the dentate gyrus. There are two positive feedback loops within the hippocampus proper, linked in a positive feedback loop. Pyramidal neurons in CA3 heavily target other CA3 pyramidal neurons. Mossy cells in the dentate gyrus target granule cells, and granule cells target mossy cells. In addition, the granule cells target CA3 pramidals. The positive feedback loops are kept in check by the granule cells also targetting inhibitory interneurons in CA3, which in turn target CA3 pyramidals. At relatively low levels of granule cell activity the excitatory connectivity predominates, but as granule cell activity increases, the inhibitory connectivity becomes predominant.
The receptive fields in the parahippocampal and perirhinal cortices correspond with groups of cortical columns that expanded their receptive fields at similar times in the past. Receptive fields in the entorhinal cortex correspond with large groups of cortical columns that expanded their receptive fields at similar times in the past. Receptive fields in CA1, CA3 and granule cells in the dentate gyrus correspond with even larger groups of cortical columns that expanded their receptive fields at similar times in the past. Granule cells target CA3 pyramidal neurons with similar receptive fields, both directly and via interneurons. CA3 pyramidals target CA1 pyramidals with similar receptive fields, and CA1 pyramidals target layer V/VI of the entorhinal cortex (via an intermediate structure called the subiculum). Layer V/VI entorhinal activity targets layer V/VI of the cortex directly or via the parahippocampal or perirhinal cortices.
The selection mechanism can be understood as follows. Suppose that the brain is experiencing a very familiar situation. There will be strong activity in all participating cortical areas. As a result there will be strong activity in all the granule cells. This strong granule cell activity means that inhibition of CA3 predominates over excitation, and CA3 will be shut down. With CA3 shut down, there is no activity in CA1 and no cortical receptive field expansions are driven. However, suppose that there is novelty in the current situation. Because of the novelty, activity in some cortical areas will be lower. As a result, the activity of granule cells with receptive fields corresponding with those areas will be lower. This will allow the activity of CA3 pyramidals with receptive fields corresponding with those areas to build up. As CA3 activity builds, granule cell activity will increase and further increase in CA3 pyramidals will be blocked. The net effect is activity in CA3 proportional to the degree of novelty in the current situation, with the CA3 activity corresponding with cortical areas requiring receptive field expansion.
CA3 activity drives activity in CA1. The CA1 receptive fields are slightly more focussed on the optimum groups of receptive fields because layer II entorhinal outputs target CA3 but layer III outputs target CA1. CA1 outputs indicate the very large groups of cortical columns appropriate for receptive field expansion. They drive activity in the deep layers of the entorhinal cortex in neurons corresponding with large groups of cortical columns that are members of a number of the very large groups selected by the hippocampus proper. Entorhinal cortex outputs drive activity in the deep layers of the perirhinal and parahippocampal cortices corresponding with groups of columns that are members of a number of the large groups selected in the entorhinalcortex. Outputs from the hippocampal cortices drive activity in cortical columns that are members of a number of the selected groups.
CORTICAL CHANGE MANAGEMENT BY THE HIPPOCAMPAL SYSTEM The main reason for the existence of cortical columns in the cortex is to provide the information needed to manage changes to cortical receptive fields in such a way that side effects on the value of recommendation strengths established by previous learning are minimized. The hippocampal system uses information about the internal activity of columns to determine when receptive field expansions are needed and which receptive fields should be expanded. Outputs from the hippocampal system drive the needed expansions.
The excitatory feedback loops in the hippocampus proper that are needed to identify the most appropriate receptive fields for expansion mean that there is a tendency for hippocampal problems to result in waves of irregular electrical activity across the cortex. For this reason, hippocampal problems often result in epileptic episodes.
Episodic memory is the ability to recall individual events from the past, especially events with a degree of novelty. The receptive fields in the hippocampal system recording groups of cortical columns that expanded at the same time in the past have some similarities with the type of information needed for episodic recall, and for that reason the hippocampal system is also involved in such recalls.